
the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 03 Jun 2021
| 03 Jun 2021
An investigation of the effects of BMPR1B, BMP15, and GDF9 genes on litter size in Ramlıç and Dağlıç sheep
Koray Çelikeloğlu
Mustafa Tekerli
Metin Erdoğan
Serdar Koçak
Özlem Hacan
Zehra Bozkurt
This study was carried out to determine the presence of polymorphisms in genes affecting litter size. The SNPs in bone morphogenetic protein receptor type 1B (BMPR1B), bone morphogenetic protein 15 (BMP15), and growth differentiation factor 9 (GDF9) genes were detected in 60 uniparous and 60 multiparous ewes from Ramlıç and Dağlıç breeds. The ewes are maintained in nine public herds at the breeding station of the Afyonkarahisar Sheep and Goats Breeders' Association and lambed in two consecutive breeding seasons. PCR and DNA sequencing analyses were conducted, and 36, 4, and 11 SNPs in Ramlıç and 40, 3, and 11 SNPs in Dağlıç were detected in BMPR1B, BMP15, and GDF9 genes, respectively. A total of 16 SNPs in Ramlıç and 10 SNPs in Dağlıç breeds for three genes were found to be significant (P<0.05). The resulting analyses showed that four SNPs (g.49496G>A, c.1658A>C, c.2037C>T, c.2053C>T) of the BMPR1B gene and one deletion mutation (c.28_30delCTT) in the BMP15 gene of the Ramlıç breed as well as five SNPs (c.1487C>A, c.2492C>T, c.2523G>A, c.2880A>G, and c.2763G>A) of the BMPR1B gene of the Dağlıç breed have significant positive regression coefficients in the desired direction of the rare allele. The observed mutations have potential to be used as genetic markers in the selection of prolific animals for both breeds.
- Article
(168 KB) - Full-text XML
-
Supplement
(212 KB) - BibTeX
- EndNote





Crossbreeding with various foreign breeds is performed worldwide to increase the yield of native sheep breeds. Ramlıç sheep were developed by crossbreeding American-originated Rambouillet rams with native Dağlıç ewes (Akçapınar, 2000). The Dağlıç sheep is one of the oldest autochthonous breeds in Asia Minor and has existed since the ancient era. The abilities of Dağlıç to resist the harsh environment, as well as its superior meat and fleece characteristics of the Rambouillet, were combined with the composite breed Ramlıç, carrying 65 %–70 % Rambouillet and 30 %–35 % Dağlıç genotypes. The average lamb birth weight and weaning weight in Ramlıç and Dağlıç breeds were reported as 4.26 and 30.39 as well as 3.54 and 23.25 kg, respectively. These figures hit 80 and 123 as well as 46 and 53 kg in mature ewes and rams in public herds. In addition, the litter size was reported between 1.23 and 1.56 and between 1.02 and 1.04 per ewe, respectively (Akçapınar, 2000; Koçak et al., 2016; Soysal, 2010; Tekerli et al., 2016; Yalçın, 1982).
The profitability of meat type from sheep breeding depends on the weight and litter size. Therefore, selection usually concentrates on these characteristics (Kumm, 2009; Notter, 2008; Tosh and Kemp, 1994). Due to low heritabilities of fertility-related traits, using molecular techniques and combining the results with numerical data is becoming important in this field (Pramod et al., 2013). The fact that economic conditions require rapid genetic progress makes it necessary to choose viable animals. In order to detect genetic variation in animal populations, DNA-level studies would give more reliable results, and many methods such as SNP, microsatellites, RFLP, and microarrays are used for these purposes. It was reported that SNPs have a greater potential than the others for realizing genetic progress (Pramod et al., 2013).
Genetic mutations in fecundity genes can change ovulation rate and litter size (Karsli et al., 2012; Mahdavi et al., 2014). Some of these are BMPR1B, BMP15, and GDF9 genes belonging to the super family of transforming growth factor β (TGFβ) obtained from the ovary (Drouilhet et al., 2013). These three genes, which are closely related to each other, have a significant impact on the ovulation rate and litter size (Pramod et al., 2013). These multiple birth genes belonging to the TGFβ superfamily are dominant, and the mutation of a single allele is sufficient to change the ovulation rate and litter size (Davis, 2005; Heaton et al., 2017). BMPR1B is on chromosome 6 and encodes the bone morphogenetic protein 1B receptor in the ovary (Davis, 2005). The mutation (FecB or Booroola) in this gene has a role in multiple birth. Australian Booroola Merino is one of the prolific breeds carrying this mutation. It is developed by crossing Merino and Garole sheep (Davis et al., 1982). This type of mutation is also common to some Asian sheep breeds such as Chinese small-tailed Han and Hu sheep, Indian Garole and Kendrapada sheep, Iranian Kalehkoohi sheep, and Indonesian thin-tailed Javanese sheep (Davis et al., 2002, 2006; Fogarty, 2009; Mahdavi et al., 2014; Mulsant et al., 2001; Tang et al., 2018). BMP15, also known as the GDF9B (FecX) gene, is on the X chromosome and encodes bone morphogenetic protein 15, playing an important role in follicular development in sheep (Hanrahan et al., 2004). The same gene causing prolificacy in heterozygous Romneys is called the Inverdale gene (FecXI). The mutations of this gene identified in various breeds are called by different names such as FecXI, FecXH, FecXG, FecXB, FecXL, FecXR, FecXGr, and FecXO (Alink et al., 2005; Davis, 2005; Davis et al., 1999). The GDF9 (FecG) gene on the fifth chromosome of sheep is also known to affect litter size and ovulation rate. Eight different point mutations (G1–G8) were detected on this gene. Ewes carrying a single copy of the FecG in Cambridge and Belclare sheep breeds had a 1.4 times higher ovulation rate (Davis, 2004, 2005).
This study aimed to determine the possible associations of mutations in BMPR1B, BMP15, and GDF9 genes with the litter size in Ramlıç and Dağlıç sheep and investigate the possibilities of using these relationships in a selection program.
2.1 Localization and animal material
The study was carried out in the Emirdağ (39∘1′1′′ N, 31∘9′0′′ E, 970 m) and Bolvadin (38∘42′40′′ N, 31∘2′55′′ E, 995 m) districts of the Afyonkarahisar province in Turkey. The research was conducted in nine enterprises within the scope of the “National Community-Based Small Ruminant Improvement” project coordinated by the General Directorate of Agricultural Research and Policies of the Ministry of Agriculture and Forestry. In these farm operations, sheep were fed on a pasture basis, and the additional concentrate and roughage supply was provided in times of feed shortage. A total of 30 uniparous and 30 multiparous ewes per breed (Ramlıç and Dağlıç), lambing in two consecutive breeding seasons, were used. An ethical committee approval was obtained from the Afyon Kocatepe University Animal Experiments Local Ethics Committee with decision no. 49533702/18 dated 25 February 2016.
2.2 Sample collection and DNA isolation
Blood samples were collected from the V. jugularis of the animals and then placed into vacuum tubes with EDTA. The blood samples were brought to the laboratory in a cold chain, kept at +4 ∘C until the DNA isolation stage, and then stored at −20 ∘C in cryotubes. Genomic DNA was obtained using a commercial DNA isolation kit (Thermo K0722). DNA samples were checked for integrity with a 0.6 % agarose gel; the amount and quality were measured using spectrophotometer devices (Multiscan GO and Qubit). Subsequently, the DNA samples were adjusted as 20 ng µL−1.
2.3 PCR and DNA sequencing analysis
Primers prepared with the FastPCR 6.0 package program (Kalendar et al., 2009) given in Table 1 were used for PCR analysis. Primers were designed by using intronic parts to sequence the related exons.
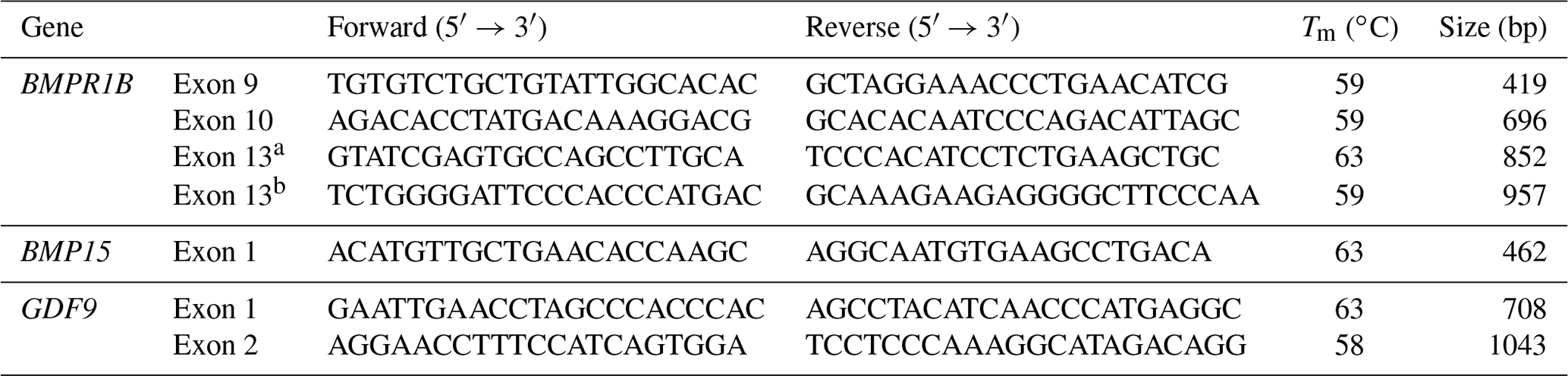
The PCR mixture of 25 µ L is comprised of 2 µ L of DNA (20 ng µ L−1), 1.25 µ L of each primer (10 pmol), 10×Q5 reaction buffer 2.5 µ L, 0.5 µ L (10 mM, Roche) of dNTP mixture, Q5 high-fidelity Taq polymerase (NEB) 0.25 µ L, and 18.5 µ L of ddH2O. The reactions were carried out on the ABI Veriti PCR instrument. The PCR apparatus was programmed at 98 ∘C for 30 s: 35 cycles in 10 s at 98 ∘C, Tm (melting temperature) 30 s, 30 s at 72 ∘C, and 10 min at 72 ∘C in the last phase. 5 µL of the PCR product, 0.5 µL Exonuclease-I (Thermo, EN0582), and 1.0 µL FastAP (Thermo, EF0652) were added into the PCR tube to clean the PCR products. The prepared mixture was placed into the PCR and adjusted to 15 min at 37 ∘C and 15 min at 85 ∘C. For DNA sequencing analysis, a total of 20 µL consisting of 2 µ L from BigDye Terminator v3.1, 11 µL of 1×sequencing buffer, 5 µL of forward or reverse primer (1 pmol), and 2 µ L of cleaned PCR product were prepared, and the PCR was programmed with the purpose of pre-denaturation at 96 ∘C for 30 s, separation at 96 ∘C for 10 s, adhesion at 54 ∘C for 15 s, and extension at 60 ∘C for 4 min. The sequence PCR products were then cleaned according to the company's protocol with BigDye XTerminator (Applied Biosystems) and placed in the DNA sequencing device (ABI 3500) after a short centrifuge. Each sample was arranged with Sequencher 5.4.6 (Gene Code Corporation, Ann Arbor, Michigan, USA) and aligned with BioEdit 7.0.9 Sequence Alignment software (Hall, 1999) after the DNA sequencing analysis. Furthermore, the nucleotide sequences determined in the BMPR1B, BMP15, and GDF9 genes were aligned with the reference sequences from the NCBI (NM_001009431.1, NM_001114767.1, and NM_001142888.2), and the sequences were translated and compared with the amino acid sequences of the references (NP_ 001009431.1, NP_001108239.1, and NP_ 001136360.2) in BioEdit.
2.4 Statistical analyses
The model for analyzing the associations between litter size and SNPs in each breed was , where the variants of the SNPs are considered covariates. In this equation Yijk indicates litter size, μ is the overall mean, H is the fixed effect of the herd, A is the random individual effect, b is the linear regression coefficient between the variants of the SNPs and litter size, and eijk is random error N (0, σ2). According to Aulchenko et al. (2007) and Khatib (2015) this model takes SNPs as the covariate. Genotypes are coded as 0, 1, and 2 for none, a single copy, and double copies according to the presence of the rare allele. A pedigree file was produced using the records of 70310 individuals. Unpublished software called Pedigri Yıldızı (Tekerli, 2018) was used to create the file. WOMBAT (Meyer and Tier, 2012) software and the run option –snapp were used in the analysis of the model, including SNP effects.
Different regions of BMPR1B, BMP15, and GDF9 genes were examined in the blood samples from Ramlıç and Dağlıç breeds. The populations were found to be in Hardy–Weinberg equilibrium (P<0.05) with a total heterozygosity of 0.1491 and 0.1638.
3.1 BMPR1B gene
As a result of PCR analysis, PCR products with the size of 419 bp (base pair) at exon 9, 696 bp at exon 10, and 1809 bp at exon 13 in the BMPR1B gene were obtained. The distributions of 36 SNPs determined in the BMPR1B gene of the Ramlıç breed at the end of the DNA sequencing analysis were as follows: four SNPs in intron 8, three SNPs in exon 9, one SNP in intron 9, two SNPs in exon 10, four SNPs in intron 10, two SNPs in intron 12, and one SNP in exon 13 and 19 in the 3′ UTR. The g.49480C>T, g.49496G>A, c.1658A>C, c.1875T>C, c.1890C>T, c.2037C>T, c.2053C>T, c.2070G>C, c.2083C>T, c.2129C>T, c.2492C>T, c.2523G>A, c.2763G>A, and c.2978C>T were statistically significant (P<0.05) SNPs detected in the 12th intron, 13th exon, and 3′ UTR regions (Table 2).
Table 2The SNPs located in the 12th intron, 13th exon, and 3′ UTR region in the BMPR1B gene of Ramlıç sheep, along with their significance levels and calculated regression coefficients.
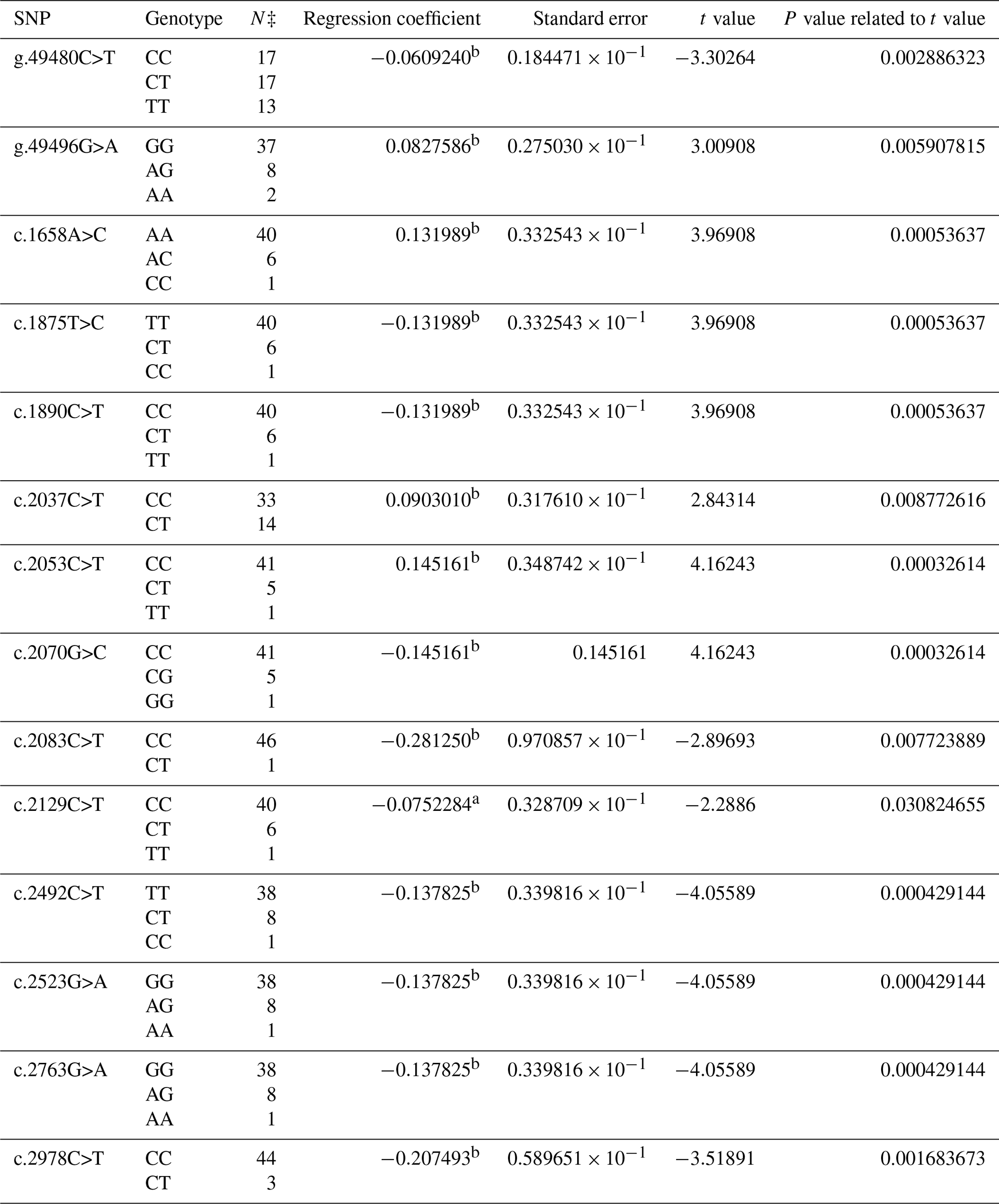
a P<0.05; b P<0.01; N‡: number of sheep carrying related genotype.
The composition of 40 SNPs determined in the BMPR1B gene of the Dağlıç breed was as follows: five SNPs in intron 8, three SNPs in exon 9, one SNP in intron 9, two SNPs in exon 10, five SNPs in intron 10, two SNPs in intron 12, and one SNP in exon 13 and 21 in the 3′ UTR. The c.1487C>A, c.1658A>C, c.1659T>A, c.1875T>C, c.1890C>T, c.2053C>T, c.2070G>C, c.2880A>G, and c.2978C>T were statistically significant (P<0.05) SNPs detected in the 13th exon and 3′ UTR regions (Table 3).
Table 3The SNPs located in the 13th exon and 3′ UTR region in the BMPR1B gene of the Dağlıç breed, along with their significance levels and calculated regression coefficients.
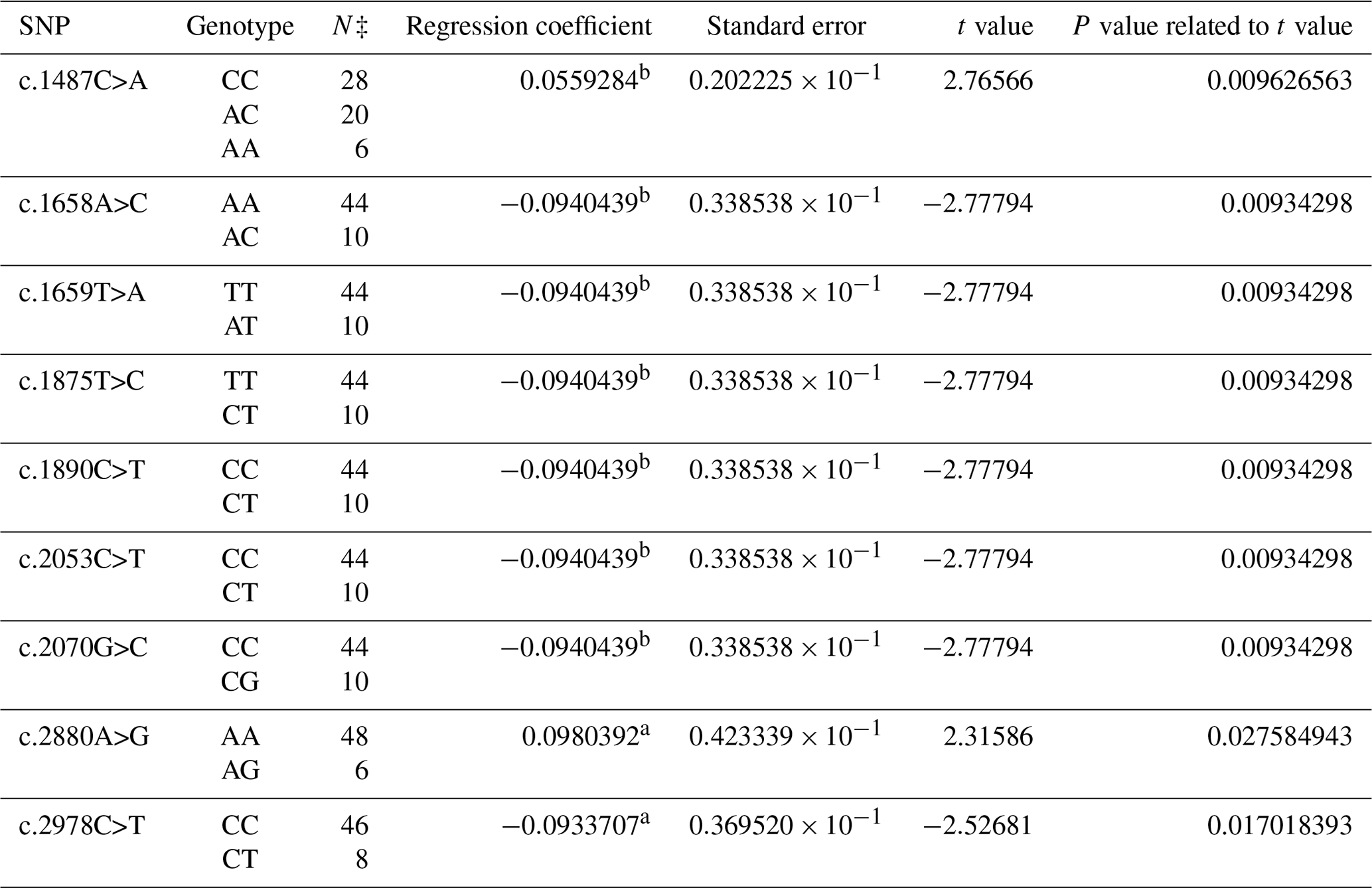
a P<0.05; b P<0.01; N‡: number of sheep carrying related genotype.
3.2 BMP15 gene
A PCR product with the size of 462 bp was obtained in the first exon of the BMP15 gene. A total of three SNPs in both breeds were found at the end of DNA sequencing analysis. Two of them were in the first exon and one of them was in the first intron of the BMP15 gene. It was determined that c.28_30delCTT and c.199C>T in the Ramlıç breed as well as g.383G>A in the Dağlıç breed were statistically significant (P<0.05) SNPs detected in the first exon and first intron (Tables 4 and 5).
Table 4The SNPs located in the first exon and intron in the BMP15 gene of the Ramlıç breed, with their significance levels and calculated regression coefficients.
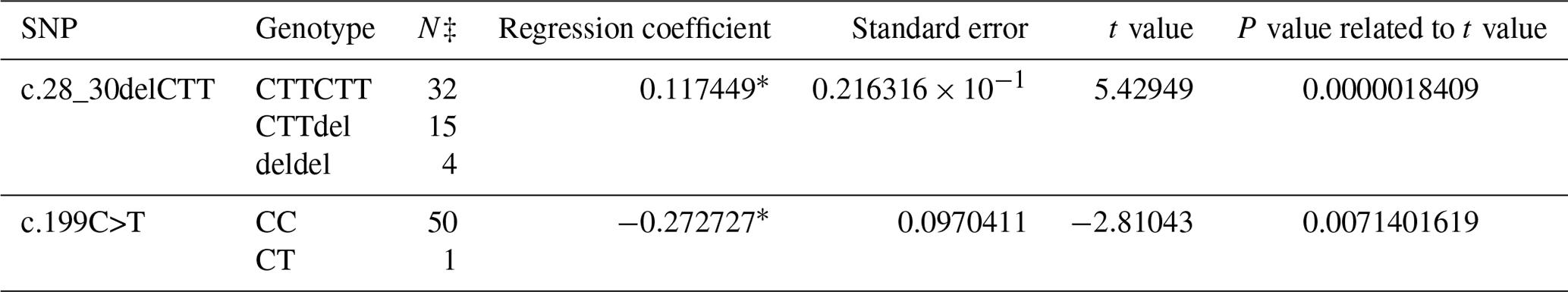
* P<0.01; N‡: number of sheep carrying related genotype.
Table 5The SNPs located in the first exon and intron in the BMP15 gene of the Dağlıç breed, with their significance levels and calculated regression coefficients.
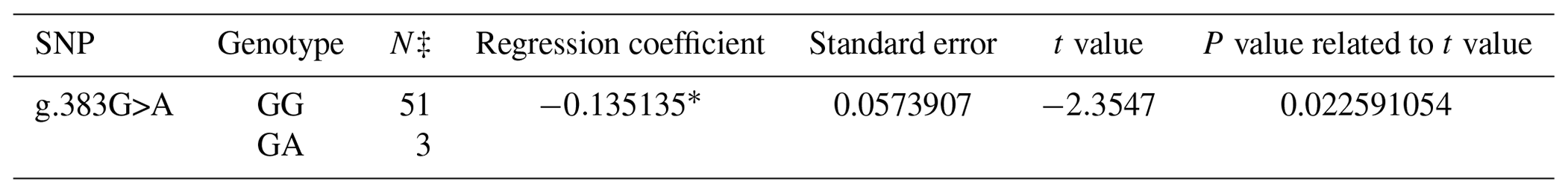
* P<0.05; N‡: number of sheep carrying related genotype.
3.3 GDF9 gene
PCR products with the size of 708 bp in exon 1 and 1043 bp in exon 2 were obtained in the GDF9 gene. At the end of the DNA sequencing analysis, a total of 11 SNPs were obtained; two of them were in exon 1, two in intron 1, and seven in exon 2 of the GDF9 gene of the Ramlıç breed. A total of 11 SNPs were also obtained in the Dağlıç breed; two were in the first exon, one in the first intron, and eight in the second exon. The regression coefficients of SNPs in this group were not statistically significant.
4.1 BMPR1B gene
The findings of this study showed that SNPs detected in the BMPR1B gene were consistent with the results in the literature for Booroola Merino, Han and Hu, Kendrapada, and Javanese sheep (Davis et al., 2002, 2006; Fogarty, 2009; Heaton et al., 2017). The effects of SNPs located in intron 8, exon and intron 9, and exon and intron 10 were not statistically significant in either breed. The non-significant results may have originated from the methods of the research, the breeds, and/or the models used in statistical evaluations. No literature studies reporting significant effects of these SNPs were encountered. FecB is a point mutation at the nucleotide position 830, which results in the replacement of the glutamine-to-arginine amino acid (Q249R) (Souza et al., 2001). This mutation was not able to be detected in the present study. However, 22 new SNPs with statistical significance were identified. The significant and positive regression coefficients for SNPs of Ramlıç (g.49496G>A, c.1658A>C, c.2037C>T and c.2053C>T) and Dağlıç (c.1487C>A, c.2492C>T, c.2523G>A, c.2880A>G, and c.2763G>A) breeds showed that the individuals carrying rare alleles have more potential for multiple births.
4.2 BMP15 gene
A total of four SNPs were found in the BMP15 gene: three in the first exon and one in the first intron. Similar results indicating that different mutations may be encountered in the gene were reported by different researchers (Davis, 2005; Demars et al., 2013; Drouilhet et al., 2013; Gürsel, 2009; Monteagudo et al., 2009) in Romney, Belclare, Cambridge, Lacaune, Rasa Aragonesa, Grivette, and Olkuska sheep. The c.28_30delCTT mutation with low frequencies detected in exon 1 of this gene of the Ramlıç breed causes the 10th amino acid (leucine) not to take part in protein synthesis. Different researchers (Demars et al., 2013; Hanrahan et al., 2004) have found similar results, determining the c.28_30delCTT mutation in Cambridge, Belclare, Olkuska, and Grivette sheep, but alleged that it was not functional in the occurrence of multiple births. However, it was highly significant (P<0.000001) in this study, and the direction of the regression line showed that the rare allele can increase the rate of multiple births. The SNP (c.199C>T) in Ramlıç and the SNP (g.383G>A) in Dağlıç breeds were also significant, but the regression coefficients were negative. Therefore, the selection for wild-type alleles will be more beneficial in sheep improvement programs. No literature studies on mutations, other than the c.28_30delCTT deletion mutation detected in the BMP15 gene, have been encountered.
4.3 GDF9 gene
A total of 13 SNPs were detected in the gene: two in the first exon, two in the first intron, and nine in the second exon in both Ramlıç and Dağlıç breeds. This finding is consistent with the results reported by different researchers (Davis, 2004, 2005; Demars et al., 2013; Hanrahan et al., 2004; Våge et al., 2013) in Cambridge, Belclare, Santa Inês, and Norway White sheep. Hanrahan et al. (2004) reported that c.260G>A, c.471C>T, c.477G>A, c.721G>A, c.978A>G, and c.994G>A point mutations were non-functional, in agreement with this study.
It has been reported in the literature that the litter size is affected by the BMPR1B, BMP15, and GDF9 genes. The effects of the first two genes were also significant for both breeds in this study. Accordingly, the regression coefficients of the SNPs for Ramlıç (g.49496G>A, c.1658A>C, c.2037C>T, and c.2053C>T) and Dağlıç breeds (c.1487C>A, c.2492C>T, c.2523G>A, c.2880A>G, and c.2763G>A) for the BMPR1B gene were statistically significant and in the desired direction of rare alleles. Significant mutation of c.28_ 30delCTT in the Ramlıç breed for BMP15 showed that the rare alleles may be functional. The SNP differences in the two breeds showed that crossbreeding (Rambouillet with Dağlıç) increased the allelic diversity in Ramlıç sheep. Polymorphisms found in both breeds would be beneficial in selection programs for producing more prolific animals.
WOMBAT software is available at http://didgeridoo.une.edu.au/km/wombat.php (last access: June 2021) (REML, 2021).
The data from this study can be obtained from the authors upon reasonable request.
The supplement related to this article is available online at: https://doi.org/10.5194/aab-64-223-2021-supplement.
KÇ, MT, and ME were responsible for the study design. KÇ, SK and ZB collected the data. KÇ, MT, ME and ÖH performed data analysis. KÇ wrote the paper. All authors read and approved the final article.
The authors declare that they have no conflict of interest.
The authors wish to thank the Scientific and Technological Research Council of Turkey (grant no. TÜBİTAK – TOVAG 116O364), the Republic of Turkey Ministry of Agriculture and Forestry General Directorate of Agricultural Research and Policies (grant no. TAGEM 03RAM2011-01 and 03DAG2011-01), and Samet Çinkaya and Mustafa Demirtaş for their assistance.
This research has been supported by the Scientific and Technological Research Council of Turkey (grant no. TÜBİTAK – TOVAG 116O364) and the Republic of Turkey Ministry of Agriculture and Forestry General Directorate of Agricultural Research and Policies (grant no. TAGEM 03RAM2011-01 and 03DAG2011-01).
This paper was edited by Steffen Maak and reviewed by Fatih Yildirim and two anonymous referees.
Akçapınar, H.: Koyun Yetiştiriciliği, Medisan yayınevi, Ankara, 2000.
Alink, F. M., Mylne, M. J. A., Watt, R. G., Kenyon, P., Wood, M. J., and McEvoy, T. G.: Inverdale fecundity gene (FecX I) influences twin ovulation incidence in pubertal ewe lambs from Texel sires and Cheviot or Scottish Blackface dams, Proceedings of the British Society of Animal Science, 2005, 55–55, https://doi.org/10.1017/s1752756200009662, 2005.
Aulchenko, Y. S., De Koning, D. J., and Haley, C.: Genomewide rapid association using mixed model and regression: A fast and simple method for genomewide pedigree-based quantitative trait loci association analysis, Genetics, 177, 577–585, https://doi.org/10.1534/genetics.107.075614, 2007.
Davis, G. H.: Fecundity genes in sheep, Anim. Reprod. Sci., 82, 247–253,https://doi.org/10.1016/j.anireprosci.2004.04.001, 2004.
Davis, G. H.: Major genes affecting ovulation rate in sheep, Genet. Sel. Evol., 37, 11–23, https://doi.org/10.1051/gse:2004026, 2005.
Davis, G. H., Montgomery, G. W., Allison, A. J., Kelly, R. W., and Bray, A. R.: Segregation of a major gene influencing fecundity in progeny of booroola sheep, New Zeal. J. Agr. Res., 25, 525–529, https://doi.org/10.1080/00288233.1982.10425216, 1982.
Davis, G. H., Dodds, K. G., and Bruce, G. D.: Combined Effect of the Inverdale and Booroola Prolificacy Genes on Ovulation Rate in Sheep, Proceedings of the Association for the Advancement of Animal Breeding and Genetics, 3, 0–12, 1999.
Davis, G. H., Galloway, S. M., Ross, I. K., Gregan, S. M., Ward, J., Nimbkar, B. V., Ghalsasi, P. M., Nimbkar, C., Gray, G. D., Subandriyo, Inounu, I., Tiesnamurti, B., Martyniuk, E., Eythorsdottir, E., Mulsant, P., Lecerf, F., Hanrahan, J. P., Bradford, G. E., and Wilson, T.: DNA tests in prolific sheep from eight countries provide new evidence on origin of the Booroola (FecB) mutation, Biol. Reprod., 66, 1869–1874, https://doi.org/10.1095/biolreprod66.6.1869, 2002.
Davis, G. H., Balakrishnan, L., Ross, I. K., Wilson, T., Galloway, S. M., Lumsden, B. M., Hanrahan, J. P., Mullen, M., Mao, X. Z., Wang, G. L., Zhao, Z. S., Zeng, Y. Q., Robinson, J. J., Mavrogenis, A. P., Papachristoforou, C., Peter, C., Baumung, R., Cardyn, P., Boujenane, I., Cockett, N. E., Eythorsdottir, E., Arranz, J. J., and Notter, D. R.: Investigation of the Booroola (FecB) and Inverdale (FecXI) mutations in 21 prolific breeds and strains of sheep sampled in 13 countries, Anim. Reprod. Sci., 92, 87–96, https://doi.org/10.1016/j.anireprosci.2005.06.001, 2006.
Demars, J., Fabre, S., Sarry, J., Rossetti, R., Gilbert, H., Persani, L., Tosser-Klopp, G., Mulsant, P., Nowak, Z., Drobik, W., Martyniuk, E., and Bodin, L.: Genome-Wide Association Studies Identify Two Novel BMP15 Mutations Responsible for an Atypical Hyperprolificacy Phenotype in Sheep, PLoS Genet., 9, 1–13, https://doi.org/10.1371/journal.pgen.1003482, 2013.
Drouilhet, L., Mansanet, C., Sarry, J., Tabet, K., Bardou, P., Woloszyn, F., Lluch, J., Harichaux, G., Viguié, C., Monniaux, D., Bodin, L., Mulsant, P., and Fabre, S.: The Highly Prolific Phenotype of Lacaune Sheep Is Associated with an Ectopic Expression of the B4GALNT2 Gene within the Ovary, PLoS Genet., 9, 1–12, https://doi.org/10.1371/journal.pgen.1003809, 2013.
Fogarty, N. M.: A review of the effects of the Booroola gene (FecB) on sheep production, Small Ruminant. Res., 85, 75–84, https://doi.org/10.1016/j.smallrumres.2009.08.003, 2009.
Gürsel, F. E.: Mutations in BMP-1B, BMP-15 and GDF-9 genes and their effects on fecundity and ovulation rate in sheep., J. Fac. Vet. Med. Istanbul Univ., 32, 57–66, 2009.
Hall, T. A.: BIOEDIT: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT, Nucleic Acid S., 41, 95–98, 1999.
Hanrahan, J. P., Gregan, S. M., Mulsant, P., Mullen, M., Davis, G. H., Powell, R., and Galloway, S. M.: Mutations in the Genes for Oocyte-Derived Growth Factors GDF9 and BMP15 Are Associated with Both Increased Ovulation Rate and Sterility in Cambridge and Belclare Sheep (Ovis aries), Biol. Reprod., 70, 900–909, https://doi.org/10.1095/biolreprod.103.023093, 2004.
Heaton, M. P., Smith, T. P. L., Freking, B. A., Workman, A. M., Bennett, G. L., Carnahan, J. K., and Kalbfleisch, T. S.: Using sheep genomes from diverse U. S. breeds to identify missense variants in genes affecting fecundity, F1000Research, 6, 1303, https://doi.org/10.12688/f1000research.12216.1, 2017.
Kalendar, R., Lee, D., and Schulman, A.: FastPCR software for PCR primer and probe design and repeat search, Genes, Genomes and Genomics, 3, 1–14, 2009.
Karsli, T., Şahin, E., Argun Karsli, B., Alkan, S., and Soner Balcioğlu, M.: An investigation of mutations (FecX G, FecX I, FecX H, FecX B) on BMP-15 gene in some local sheep breeds raised in Turkey, Journal of the Faculty of Agriculture of Akdeniz University, 25, 29–33, 2012.
Khatib, H.: Molecular and quantitative animal genetics, John Wiley & Sons., New York, USA, 2015.
Koçak, S., Tekerli, M., Erdoğan, M., Çelikeloğlu, K., Bozkurt, Z., and Hacan, Ö.: Halk Elinde Küçükbaş Hayvan IslahıÜlkesel Projesi, Dağlıç Koyunlarının Halk Elinde IslahıProjesi Kesin Raporu, Afyonkarahisar, 2016.
Kumm, K. I.: Profitable Swedish lamb production by economies of scale, Small Ruminant. Res., 81, 63–69, https://doi.org/10.1016/j.smallrumres.2008.11.006, 2009.
Mahdavi, M., Nanekarani, S., and Hosseini, S. D.: Mutation in BMPR-IB gene is associated with litter size in Iranian Kalehkoohi sheep, Anim. Reprod. Sci., 147, 93–98, https://doi.org/10.1016/j.anireprosci.2014.04.003, 2014.
Meyer, K. and Tier, B.: “SNP Snappy”: A strategy for fast genome-wide association studies fitting a full mixed model, Genetics, 190, 275–277, https://doi.org/10.1534/genetics.111.134841, 2012.
Monteagudo, L. V., Ponz, R., Tejedor, M. T., Laviña, A., and Sierra, I.: A 17 bp deletion in the Bone Morphogenetic Protein 15 (BMP15) gene is associated to increased prolificacy in the Rasa Aragonesa sheep breed, Anim. Reprod. Sci., 110, 139–146, https://doi.org/10.1016/j.anireprosci.2008.01.005, 2009.
Mulsant, P., Lecerf, F., Fabre, S., Schibler, L., Monget, P., Lanneluc, I., Pisselet, C., Riquet, J., Monniaux, D., Callebaut, I., Cribiu, E., Thimonier, J., Teyssier, J., Bodin, L., Cognié, Y., Chitour, N., and Elsen, J. M.: Mutation in bone morphogenetic protein receptor-IB is associated with increased ovulation rate in Booroola Mérino ewes, Proc. Natl. Acad. Sci. USA, 98, 5104–5109, https://doi.org/10.1073/pnas.091577598, 2001.
Notter, D. R.: Genetic Aspects of Reproduction in Sheep, Reprod. Domest. Anim., 43, 122–128, https://doi.org/10.1111/j.1439-0531.2008.01151.x, 2008.
Pramod, R. K., Sharma, S. K., Kumar, R., and Rajan, A.: Genetics of ovulation rate in farm animals, Veterinary World, 6, 833–838, https://doi.org/10.14202/vetworld.2013.833-838, 2013.
REML: WOMBAT, available at: http://didgeridoo.une.edu.au/km/wombat.php, last access: June 2021.
Souza, C. J. H., MacDougall, C., Campbell, B. K., McNeilly, A. S., and Baird, D. T.: The Booroola (FecB) phenotype is associated with a mutation in the bone morphogenetic receptor type 1 B (BMPR1B) gene, J. Endocrinol., 169, 1–6, https://doi.org/10.1677/joe.0.169R001, 2001.
Soysal, M. İ.: Türkiye Yerli Evcil Hayvan Genetik Kaynakları, Lecture notes, Namık Kemal University, Tekirdağ, 2010.
Tang, J., Hu, W., Di, R., Liu, Q., Wang, X., Zhang, X., Zhang, J., and Chu, M.: Expression analysis of the prolific candidate genes, bmpr1b, bmp15, and gdf9 in small tail han ewes with three fecundity (Fecb gene) genotypes, Animals, 8, 166, https://doi.org/10.3390/ani8100166, 2018.
Tekerli, M.: Pedigri Yıldızı; Unpublished Excel based pedigree software, 2018.
Tekerli, M., Erdoğan, M., Koçak, S., Çelikeloğlu, K., Bozkurt, Z., and Hacan, Ö.: Halk Elinde Küçükbaş Hayvan IslahıÜlkesel Projesi, Ramlıç Koyunlarının Halk Elinde IslahıProjesi Kesin Raporu, Afyonkarahisar, 2016.
Tosh, J. J. and Kemp, R. A.: Estimation of variance components for lamb weights in three sheep populations, J. Anim. Sci., 72, 1184–1190, https://doi.org/10.2527/1994.7251184x, 1994.
Våge, D. I., Husdal, M., Kent, M. P., Klemetsdal, G., and Boman, I. A.: A missense mutation in growth differentiation factor 9 (GDF9) is strongly associated with litter size in sheep, BMC Genet., 14, 1–8, https://doi.org/10.1186/1471-2156-14-1, 2013.
Yalçın, B. C.: Rambouillet x Dağlıç melezlemesiyle geliştirilmiş et yapağıyönlü yeni bir koyun tipi, J. Fac. Vet. Med. Istanbul Univ., 8, 5–15, 1982.