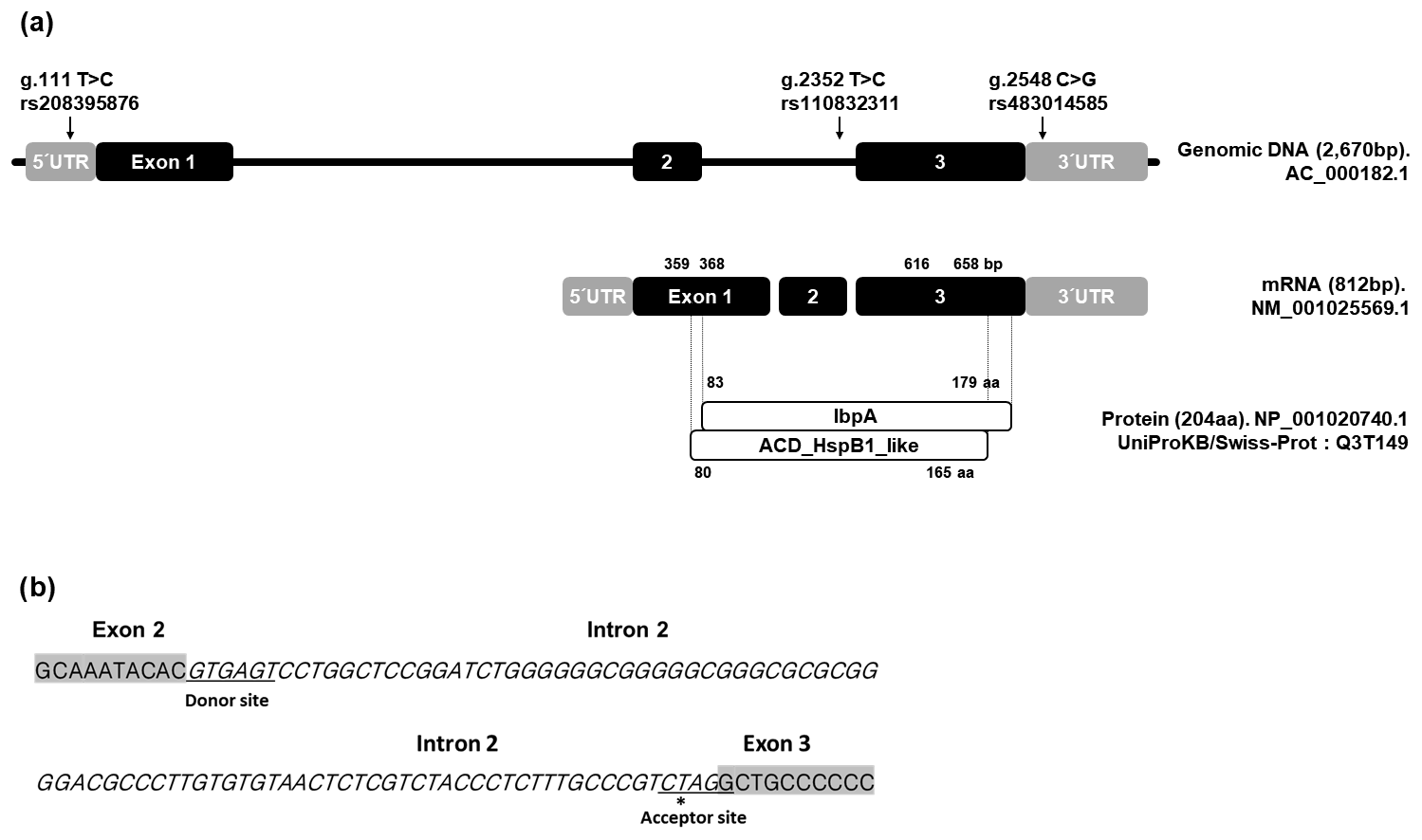
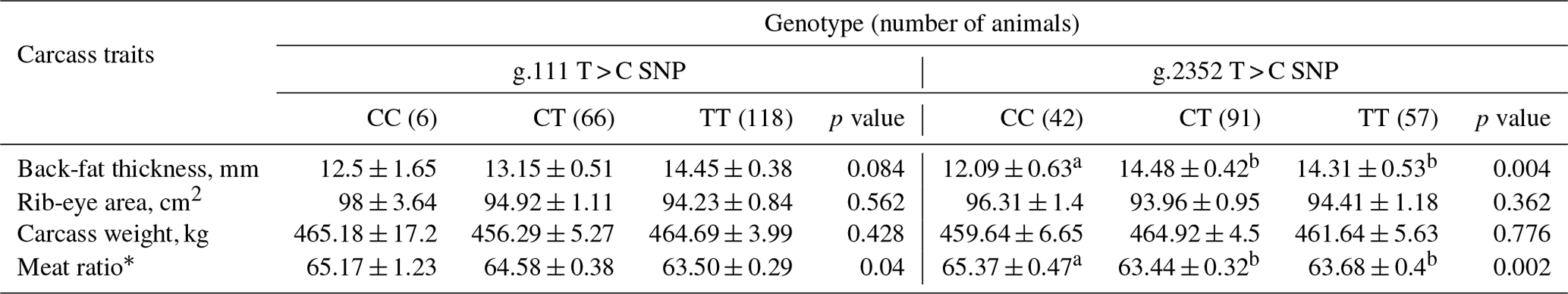
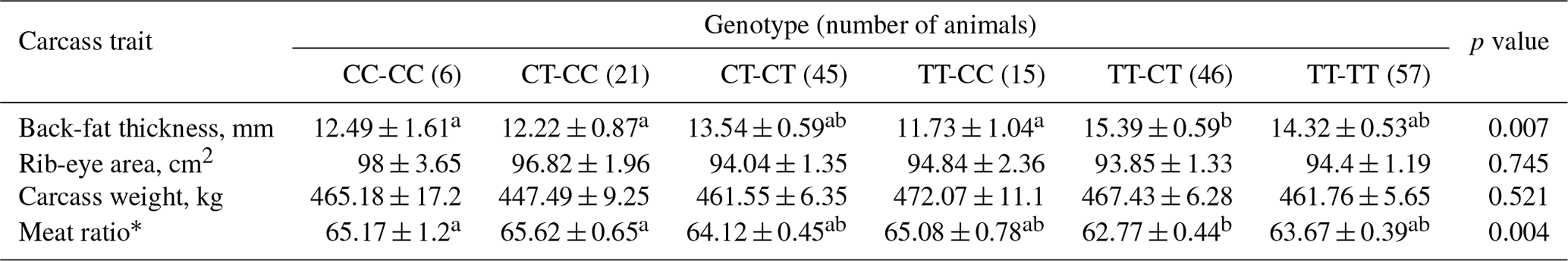
Arrigo, A.-P.: Mammalian HspB1 (Hsp27) is a molecular sensor linked to the physiology and environment of the cell, Cell Stress Chaperon., 22,
517–529, 2017.
Charge, S. B. and Rudnicki, M. A.: Cellular and molecular regulation of
muscle regeneration, Physiol. Rev., 84, 209–238, 2004.
Dubińska-Magiera, M., Jabłońska, J., Saczko, J., Kulbacka, J.,
Jagla, T., and Daczewska, M.: Contribution of small heat shock proteins to
muscle development and function, FEBS Lett., 588, 517–530, 2014.
Hamelin, M., Sayd, T., Chambon, C., Bouix, J., Bibé, B., Milenkovic, D.,
Levéziel, H., Georges, M., Clop, A., and Marinova, P.: Proteomic
analysis of ovine muscle hypertrophy, J. Animal Sci., 84,
3266–3276, 2006.
He, H., Chen, S., Liang, W., and Liu, X.: Genome-wide proteomics analysis on
longissimus muscles in Qinchuan beef cattle, Anim. Genet., 48, 131–140,
2017.
Jung, U., Kim, M., Wang, T., Lee, J., Jeon, S., Jo, N., Kim, W., Baik, M.,
and Lee, H.: Upregulated heat shock protein beta-1 associated with caloric
restriction and high feed efficiency in longissimus dorsi muscle of steer,
Livest. Sci., 202, 109–114, 2017.
Kim, N.-K., Lim, D., Lee, S.-H., Cho, Y.-M., Park, E.-W., Lee, C.-S., Shin,
B.-S., Kim, T.-H., and Yoon, D.: Heat shock protein B1 and its regulator
genes are negatively correlated with intramuscular fat content in the
Longissimus Thoracis muscle of Hanwoo (Korean cattle) steers, J.
Agr. Food Chem., 59, 5657–5664, 2011.
Kim, Y.-S., Lee, J.-S., Lee, Y., Kim, W.-S., Peng, D.-Q., Bae, M.-H., Jo,
Y.-H., Baik, M., and Lee, H.-G.: Effect of glutamine on heat-shock protein
beta 1 (HSPB1) expression during myogenic differentiation in bovine embryonic
fibroblast cells, Food Sci. Biotechnol., 27, 829–835, 2018.
Lee, Y.-S., Oh, D., Kim, J.-J., Lee, J.-H., Park, H.-S., and Yeo, J.-S.: A
single nucleotide polymorphism in LOC534614 as an unknown gene associated
with body weight and cold carcass weight in Hanwoo (Korean cattle),
Asian-Austr. J. Animal Sci., 23, 1543–1551, 2010.
Oh, D.-y., Nam, I., Hwang, S., Kong, H., Lee, H., Ha, J., Baik, M., Oh, M.
H., Kim, S., and Han, K.: In vivo evidence on the functional variation
within fatty acid synthase gene associated with lipid metabolism in bovine
longissimus dorsi muscle tissue, Genes Genom., 40, 289–294, 2018.
Perng, M. D., Cairns, L., van den IJssel, P., Prescott, A., Hutcheson, A.
M., and Quinlan, R. A.: Intermediate filament interactions can be altered by
HSP27 and alphaB-crystallin, J. Cell Sci., 112, 2099–2112, 1999.
Rajesh, R. V., Kim, S.-K., Park, M.-R., Nam, J.-S., Kim, N.-K., Kwon, S.,
Yoon, D., Kim, T.-H., and Lee, H.-J.: Proteomic Functional Characterization
of Bovine Stromal Vascular Cells from Omental, Subcutaneous and
Intramuscular Adipose Depots, Asian Austral. J. Anim.,
24, 110–124, 2010.
Sugiyama, Y., Suzuki, A., Kishikawa, M., Akutsu, R., Hirose, T., Waye, M.
M., Tsui, S. K., Yoshida, S., and Ohno, S.: Muscle develops a specific form
of small heat shock protein complex composed of MKBP/HSPB2 and HSPB3 during
myogenic differentiation, J. Biol. Chem., 275, 1095–1104,
2000.
Sweetman, D.: The myogenic regulatory factors: critical determinants of
muscle identity in development, growth and regeneration, Intech, 31–39, 2012.
Tucker, N. R. and Shelden, E. A.: Hsp27 associates with the titin filament
system in heat-shocked zebrafish cardiomyocytes, Exp. Cell Res.,
315, 3176–3186, 2009.
Zhang, Q., Lee, H.-G., Han, J.-A., Kang, S. K., Lee, N. K., Baik, M., and
Choi, Y.-J.: Differentially expressed proteins associated with myogenesis
and adipogenesis in skeletal muscle and adipose tissue between bulls and
steers, Mol. Biol. Rep., 39, 953–960, 2012.
Zhang, Q., Lee, H.-G., Kang, S. K., Baik, M., and Choi, Y.-J.: Heat-shock
protein beta 1 regulates androgen-mediated bovine myogenesis, Biotechnol.
Lett., 36, 1225–1231, 2014.