
the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 17 Mar 2022
| 17 Mar 2022
Comparison of different models for estimation of direct and maternal genetic parameters on body weights in Awassi sheep
Hatice Hızlı
Çiğdem Takma
Ertan Yazgan
The present study was conducted to estimate the (co)variance components for birth and weaning weight (BW and WW) in 8142 Awassi sheep between 2015 and 2017. Estimates were calculated with single-trait analysis by the average information restricted maximum likelihood (AI-REML) method, using a derivative-free algorithm by fitting six different univariate animal models. The negative of the log-likelihood function (LogL), Akaike information criterion (AIC), and Bayesian information criterion (BIC) tests were used for selecting the best fitted model. In addition, the goodness of fit between the two models was compared with the likelihood ratio test (LRT). Depending on the models, and ranged from 0.230 to 0.240 and 0.015 to 0.033 for BW, and 0.108 to 0.168 and 0.024 to 0.081 for WW, respectively. Model 3 for BW and Model 2 for WW were chosen as the best models by LogL comparison criteria. According to the LRT ratio test Model 2, Model 3, and Model 4 for BW and Model 2, Model 3, Model 4, Model 5, and Model 6 for WW were significant (p<0.05). Including maternal genetic or maternal permanent environmental effects in these models was found to be significant in terms of parameter estimates.
- Article
(154 KB) - Full-text XML
-
Supplement
(481 KB) - BibTeX
- EndNote





For maximizing the expected yield in livestock, the first condition is to apply selection programs in which the genetic parameters must be accurately estimated (Falconer and Mackay, 1996). Awassi sheep is one of the most common breeds grown in Turkey (TUIK, 2021). In addition, it is recognized as an important genetic resource for the sheep industry in more than 30 countries in the Middle East and all over the world (Galal et al., 2008). Consumers in the Middle East prefer Awassi mutton due to its taste. Falconer and Mackay (1996) have reported that many nongenetic factors play an important role in different quantitative characteristics. Birth weight (BW) and weaning weight (WW), which are quantitative characters, are important selection criteria for prenatal and postnatal growth performance in livestock breeding. In sheep breeding, the first 3-month weights during weaning of sheep are important production characteristics (Saatçi et al., 1999). Many researchers reported significant effects of lambing year, sex, and maternal age on BW and WW (Jawasreh and Khasawneh, 2007; Jawasreh et al., 2009; Aktaş and Doğan, 2014).
The yields to be improved by selection are affected by the genetic performance of the individual and environmental factors. In addition to reducing the effects of environmental factors to increase the degree of accuracy in selection, accurate estimation of the components that make up the genetic structure is required. In farm animals with long maternal lifespans, such as sheep, the phenotype of the offspring is determined not only by their genetic potential and environment, but also by the environment provided by their dams. The maternal effect on the offspring has been known since the beginning of animal breeding, and therefore it has been studied increasingly with developed software programs in recent years.
Direct additive, maternal additive, and maternal permanent environmental variances are considered important factors that determine the lamb's genetic gain and therefore performance (Willham, 1972; Meyer, 1992; Meyer and Kirkpatrick, 2005). In this connection, several studies have been done on heritability estimates and its constituent elements (Miraei-Ashtiani et al., 2007; Jafaroghli et al., 2010; Mohammadi et al., 2013a, b; Supakorn et al., 2013).
The aims of this study were to estimate the variance and covariance components, to compare different models for the BW–WW measurements of the Awassi lambs, and to reveal the importance of including maternal effects to design true breeding programs for genotypic selection of Awassi lambs. For this purpose, direct additive, maternal genetic and the maternal permanent environmental effects were calculated by applying six animal models.
2.1 Location and flock management
The study was carried out in Osmaniye province, districts, and villages between – N, – E, in a wide area covering an altitude of 2285 to 2400 m. Although the climate in the region differs in mountainous and lowland areas, it has the characteristic of the Mediterranean climate. In general, summers are hot and dry, and winters are mild and rainy. Average temperature is 18.2 ∘C, and average maximum temperature is 42.8 ∘C. Rainfall is higher in the winter and autumn months compared to other months, and the annual average rainfall is 767.6 mm.
Awassi is warm and very well adopted to arid conditions; it is one of Turkey's indigenous breeds with a high ability to adapt to different environments. Awassi sheep are raised in southeastern Anatolia, and the high milk production is one of the remarkable traits of the breed. Newborn lambs were kept together with their dams for three months (90 d) until weaning age. The birth weight (BW) and weaning weight (WW) at 3 months of age were recorded as growth traits. BW was taken within 24 h of the birth of lamb, and WW was taken on 3-month weights with digital hand weighing scales (up to 10 g sensitivity). The regional breeders kept their lambs in similar feeding and management conditions. Except for the winter period, they apply a feeding with a nomadic system based on the pasture and stubble grazing. They feed their animals mainly with hay and a small amount of grain in winter.
The material of this study was obtained from the Awassi Sub-Project in which elite herds were created in Osmaniye province within the scope of the national community-based animal improvement project. The records collected from 249 rams, 2416 sheep, and 8142 lambs between 2015–2017 years were used.
2.2 Statistical analysis for growth traits
A preliminary least-squares analysis of variance was performed with a general linear model (GLM) with SPSS software (IBM, 2020) to determine the main effects that will take place in the individual animal model. The least-squares analysis showed that the effects of sex of lambs (male, female), type of birth (single, twin), year of birth (2015, 2016, 2017), and age of dam (2, 3, 4, 5, 6, 7) were found to be statistically significant (p<0.05) for BW and WW, and therefore these effects were included in the animal models. In these models sex of lambs, year of birth, and type of birth were considered as fixed effects and age of dam was used as a covariate for the estimations of BW. Also, sex of lambs, year of birth, type of birth, and age of dam were considered as fixed effects and BW of lamb was used as a covariate for the estimations of WW. The farm was not taken into account as a fixed effect in the models because the lambs that are the subject of this research were reared under similar care and feeding conditions and also the farms were located close to each other.
The estimates of variance components and genetic parameters for each trait were obtained from single-trait analysis with the average information restricted maximum likelihood (AI-REML) method, using a derivative-free algorithm by WOMBAT software (Meyer, 2012). The matrix representation of six single-trait animal models that used for genetic analysis were given as follows:
where Y is the vector of observations or records; β is the vector containing fixed effects such as sex, type of birth, year of birth, and age of dam; a, m, c, and ε are the vectors of direct additive genetic effects, maternal genetic effects, maternal permanent environmental effect of sheep and the residual, respectively. X, Za, Zm, and Zc are the incidence matrices relating observations to b, a, m, and c, respectively. A is the numerator relationship matrix, and σam is the covariance between direct additive and maternal genetic effects. The (co)variance structure of the random effects in the analysis can be described as
Assumptions for (co)variance matrix involving random effects can be described as A is the numerator relationship matrix, is the direct additive genetic variance, is the maternal additive genetic variance, σam is the direct-maternal additive genetic covariance, is the maternal permanent environmental variance, is the residual variance, and Id and In are the identity matrices of an order equal to the number of dams and records, respectively.
The total heritability () for each model was calculated as , which was defined by Willham (1972). The genetic correlation between the direct additive genetic and the maternal additive genetic effects (ram) was estimated as the ratio of the estimates of the σam to the estimates of and (Falconer and Mackay, 1996).
The starting values of variance parameters for REML iterations were obtained with ANOVA estimators (Searle et al., 2006). The most appropriate model for convergence of REML solutions was assumed when the average difference in likelihood functions in successive iterations was less than 10−8. The analysis was continued by taking the previous starting values until ensuring the estimates did not change and confirming the convergence was met (Meyer, 1991, 2007).
The log-likelihood (LogL) test, Akaike information criterion (AIC), and Bayesian information criterion (BIC) were performed to determine the best model. These likelihood-based tests are and , and where p is the number of predicted parameters, N is the sample size, and r(x) is the rank of the coefficient matrix for the fixed effects in the model (Akaike, 1973; Maniatis et al., 2013; Meyer, 1992; López-Romero and Carabano, 2003; Schwarz, 1978). The model with the highest LogL or minimum AIC and BIC value is considered the best model (Pham, 2019). Moreover, the likelihood ratio test (LRT) is a statistical test of the goodness of fit between two models; a relatively full model is compared to a reduced model to determine importance of a parameter that gives rise to increase in log-likelihood. That is, the full model must differ from the reduced model only by the addition of one or more parameters. In other words, LRT explains whether it is good to add a parameter to a model or not. The test begins with a comparison of the likelihood scores of the two models, after which follows a chi-squared distribution (χ2) with degrees of freedom equal to the difference in dimensionality of the models (Felsenstein, 1981; Huelsenbeck and Crandall, 1997). The equation for the test statistic is as follows:
In this study, LogL differences between Model 1 versus other models were examined with LRT in order to determine the importance of the maternal additive genetic and maternal permanent environmental effects.
The structure of the data set used in this study is presented in Table 1. According to descriptive statistics the coefficient of variation for WW is much lower than BW (Table 1).
Table 1The data structure of BW and WW for Awassi lambs.
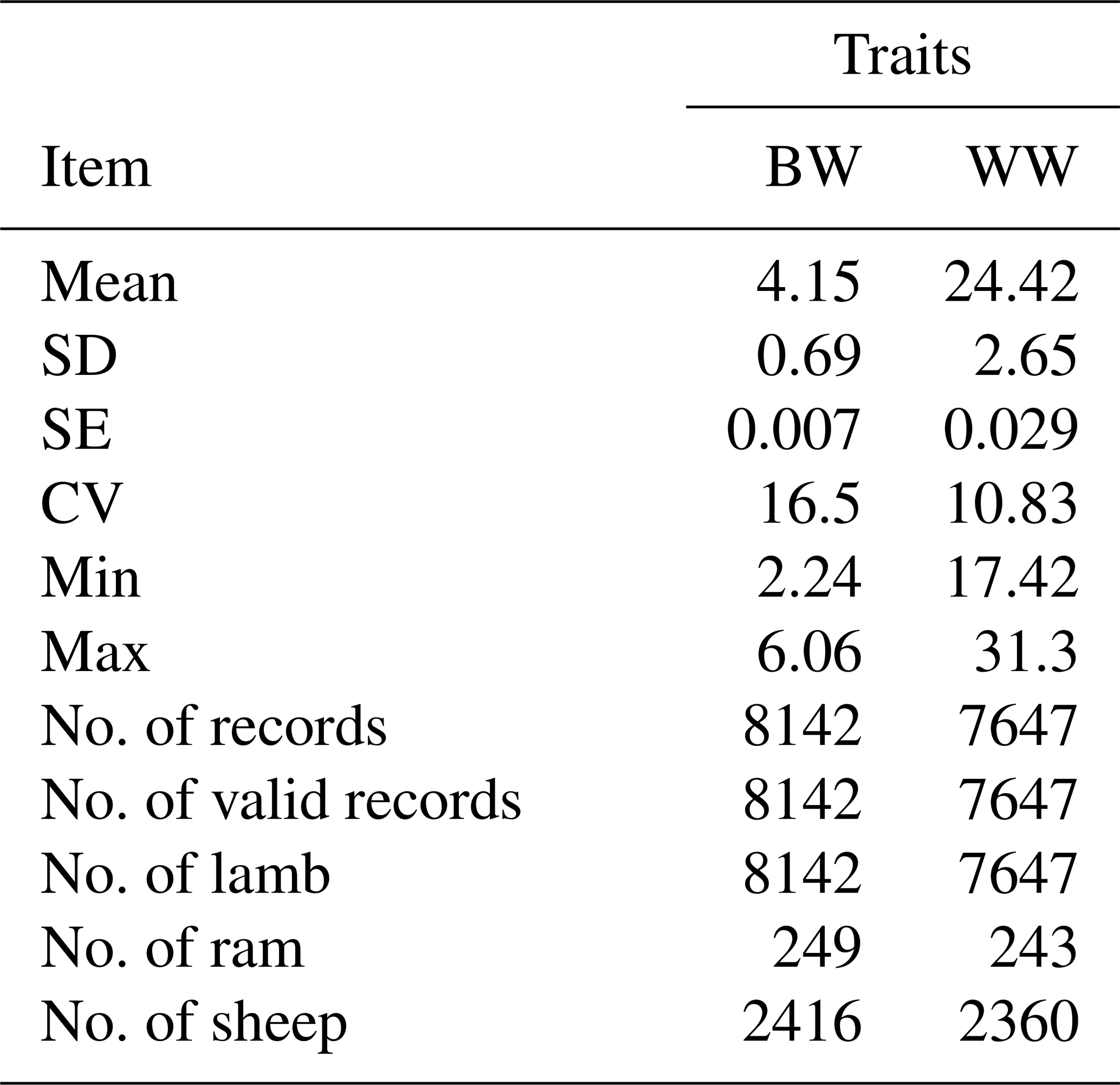
SD: standard deviation, SE: standard error, CV: coefficient of variance, Min: minimum and Max: maximum.
Least-squares means for fixed effects and standard errors for BW and WW are shown in Table 2, and they were estimated for BW and WW as 3.87±0.01 and 24.49±0.57, respectively. Sex of lambs had a significant effect on BW and WW (p<0.01). Vatankhah and Demand (2008a, b), Yavarifard et al. (2015), and Jalil-Sarghale et al. (2014) reported similar results. The result of variance analysis showed that the year of birth had significant effects on BW (p<0.01), but not significant on WW (p>0.05). Also, age of dam had a significant effect on BW and WW (p<0.001). Single-born lambs had higher body weights than twins, as expected. Similar results for age of dams and sex of offspring's growth in different species have been reported by many researchers in the literature (Gholizadeh et al., 2010; Jawasreh et al., 2018; Rashidi et al., 2011; Mohammadi et al., 2013c; Mohammadabadi and Sattayimokhtari, 2013).
Table 2The least-squares means (LSMs) and standard errors (SEs) of sex, birth type, year, and age of dam effects for Awassi lambs' BW and WW.

Different letters in the same column indicate that the difference is significant (p<0.05).
Table 3Estimates of (co)variance components and genetic parameters for BW of Awassi lambs obtained in different models (mean ± standard error). The best models according to LogL, AIC and BIC criteria are shown in italic.
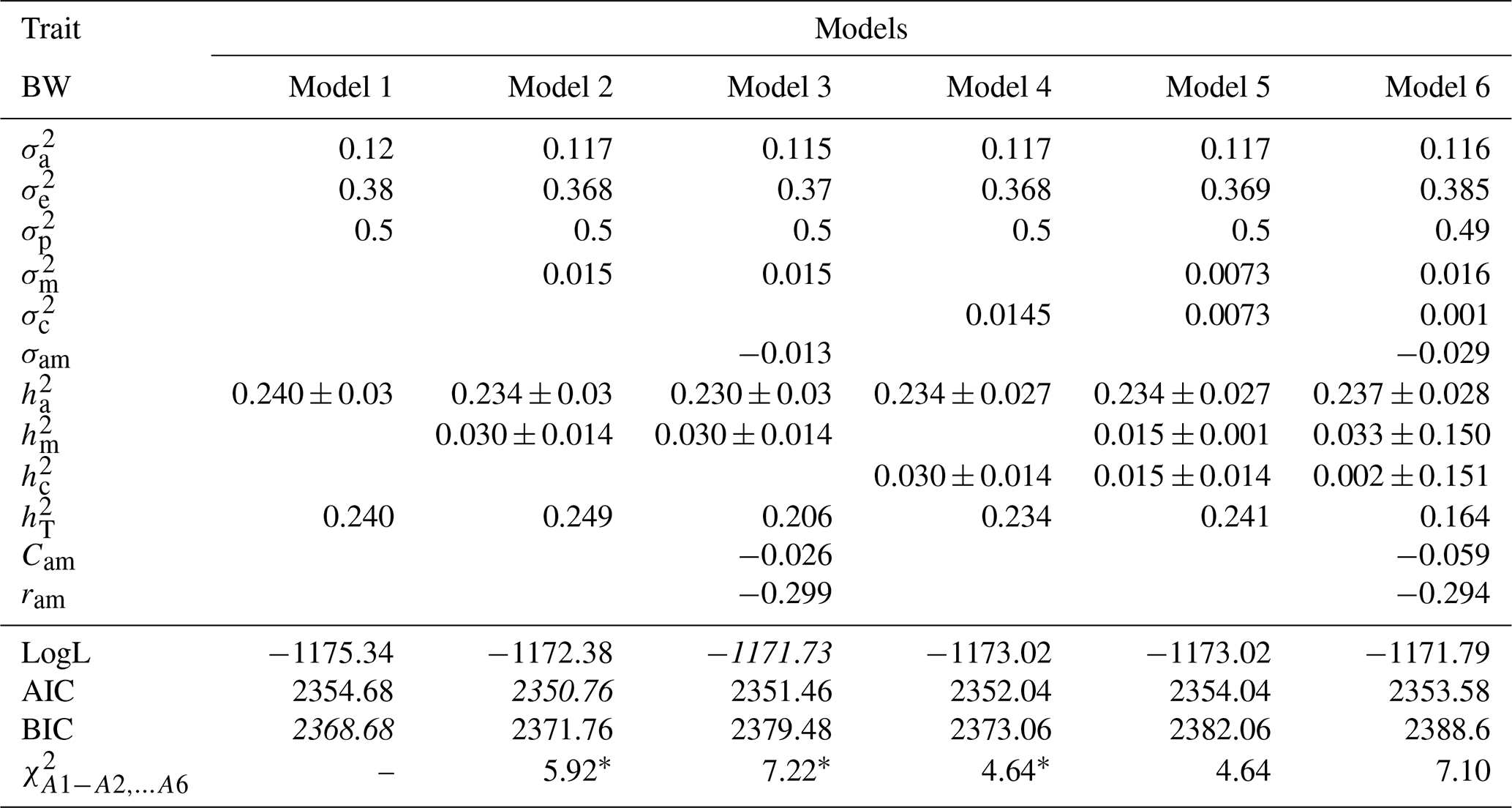
: direct additive genetic variance; : maternal additive genetic variance; σam: direct–maternal genetic covariance; : maternal permanent environmental variance; : error variance; : phenotypic variance; : direct heritability; : maternal heritability; Cam: ; ram: genetic correlation between direct and maternal effects; : maternal permanent environmental variance as proportion of phenotypic variance; : total heritability: , ; ∗: test value is significant at the 0.05 level.
Estimates of (co)variance components and genetic parameters for BWs of Awassi lambs obtained with different models are presented in Table 3. The direct additive genetic variance was found to be lower in Models 2, 3, 4, 5, and 6 when compared with Model 1. Similar results have been reported for various goat and sheep breeds (Saatçi et al., 1999; Tesema et al., 2020; Szwaczkowski et al., 2006). Estimates of maternal permanent environmental variances in Models 4, 5, and 6 for BW were as 0.0145, 0.0073, and 0.001, respectively. The estimates of Cam, which is the ratio of covariance between additive genetic and maternal genetic variances in phenotypic variance for Models 3 and 6, were estimated as −0.026 and −0.059, respectively.
Correlations between direct additive genetic and maternal genetic effects were calculated as −0.299 and −0.294 for BW in Model 3 and 6, respectively. These similar negative estimates have been reported in many studies (Ligda et al., 2000; Zhang et al., 2009; Gowane et al., 2010; Tesema et al., 2020). Meyer (1992) and Heydarpour et al. (2008) reported that higher and negative predictions of correlations are due to the presence of true genetic antagonism between components, poor data structure with few progenies per dam, and limited information about the performance of linked offspring and dams. Therefore, both additive genetic and maternal genetic effects must be considered in selection programs to maximize genetic gain (Meyer, 1992). Gutierrez et al. (1997) also suggested that this relationship should be compensated by improving management practices and using supplementary feeding.
In addition, the most suitable model was selected in italics as the best model according to LogL, AIC, and BIC criteria for the estimation of genetic parameters for the BW and WW of Awassi lambs (Tables 3 and 4). Models 1, 2, and 3 were mentioned as the best model according to BIC, AIC, and LogL criteria, respectively. Also, Models 2, 3, and 4 were found to be more statistically significant than Model 1 according to LRT (p<0.05). The attained results point out that maternal genetic effects or maternal permanent environmental effects should be included into the models for genetic evaluation of Awassi BW.
Table 4Estimates of (co)variance components and genetic parameters for WW of Awassi lambs obtained in different models (mean ± standard error). The best models according to LogL, AIC and BIC criteria are shown in italic.
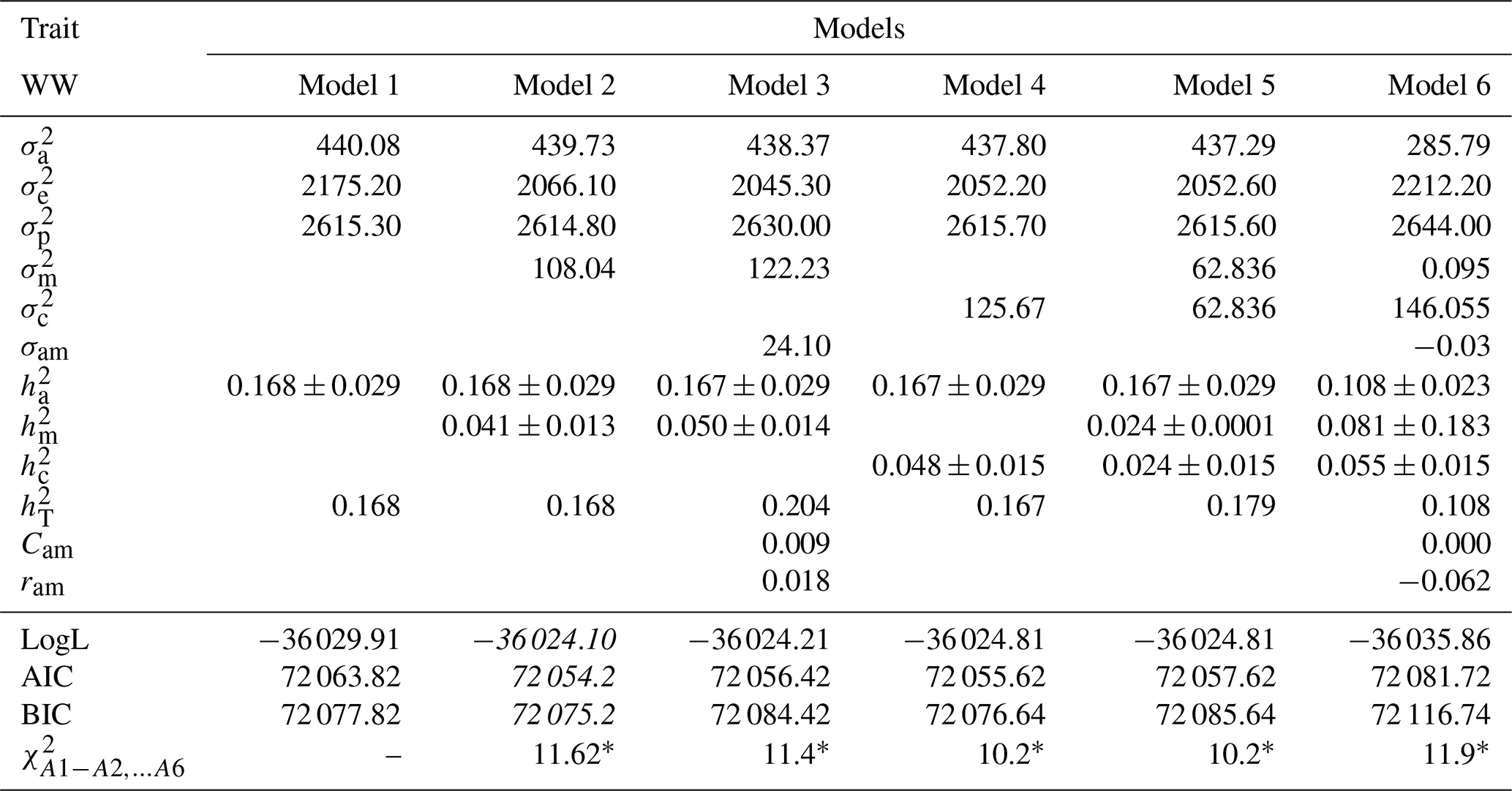
: direct additive genetic variance; : maternal additive genetic variance; σam: direct–maternal genetic covariance; : maternal permanent environmental variance; : error variance; : phenotypic variance; : direct heritability; : maternal heritability; Cam: ; ram: genetic correlation between direct and maternal effects; : maternal permanent environmental variance as proportion of phenotypic variance; : total heritability: , ∗: test value is significant at the 0.05 level.
In Table 4 the results showed that Model 2 was selected as the best model with respect to all criteria. Also, Models 2, 3, 4, 5, and 6 were found to be significant compared to Model 1 according to LRT (p<0.05). Moreover, estimates of (co)variance components and genetic parameters for WW of Awassi lambs obtained in different models are presented in Table 4. Table 4 shows that Models 5 and 6, in which maternal additive genetic and maternal permanent environmental variances are added, have lower and values than Model 1 for WW. The estimates of and in Models 2 and 3, which have maternal additive genetic variance and not maternal permanent environmental variance, were found as 439.73±0.029, 0.168±0.029, and 438.37±0.029, 0.167±0.029, respectively. In Models 2, 3, 4, 5, and 6, the maternal genetic variance and the maternal permanent environmental variance estimates of and were found to be smaller than Model 1. Cam estimates for Models 3 and 6 were estimated to be 0.009 and almost zero, respectively. Similar results have been reported by Jawasreh et al. (2018b).
In Model 4, 5, and 6 the estimates were found as 0.048±0.015, 0.024±0.015, and 0.055±0.015, respectively. Correlations between additive genetic and maternal genetic effects were obtained as 0.018 and −0.062 in Model 3 and 6 for WW, respectively.
The estimates were 0.168±0.015, 0.167±0.015, 0.204±0.015, 0.167±0.015, 0.179±0.015, and 0.108±0.015 in Models 1, 2, 3, 4, 5, and 6, respectively. The estimate was the highest in Model 3 and the lowest in Model 6. Prakash et al. (2016) reported that the maternal genetic variances are significant but are not generally considered. Therefore, the heritability estimates are biased upward and the resulting selection efficiency is reduced, but division of the (co)variances is made more accurate as it enables the estimation of the contribution of each individual effect to the overall performance of the animal.
In present study, values were estimated from 0.230 to 0.240, from 0.015 to 0.033, from 0.002 to 0.030, ram from −0.294 to −0.299, and from 0.164 to 0.240 for BW in Table 3. Similarly, , , , ram, and were estimated with values ranging from 0.108 to 0.168, 0.024–0.081, 0.024–0.055, −0.062–0.018, and 0.108–0.168, respectively for WW (Table 4). Şireli et al. (2015) found and for BW between 0.21–0.57 and 0.09–0.18, respectively, and for WW were between 0.02–0.13 and 0.03–0.17, respectively. Jawasreh et al. (2018) estimated for BW and WW as 0.30 and 0.19, respectively, and values for BW and WW were found as 0.13 and 0.03, respectively, in their study. While our estimates of (co)variance components and genetic parameters for BW and WW of Awassi lambs obtained by different models were similar with the findings of Şireli et al. (2015), it was smaller than the findings of Jawasreh et al. (2018).
Tables 3 and 4 show that the estimates of maternal genetic and maternal permanent environmental effects for WW were increased when compared with the estimates for BW. Also, Tables 3 and 4 show that the ram, , and estimates of WW were lower than the estimates for BW. The lower variance component estimates explain the magnitude and estimates for WW. While can be transferred from generation to generation, is determined by factors that cannot be transferred from generation to generation. The maternal environmental effect is the factors that are not transmitted from generation to generation and only affect the offspring, as maternal nutrition, uterine volume, litter size, and maternal care (Çelikeloğlu and Tekerli, 2014). Hanrahan (1976) and Meyer (1992) also reported that maternal effects explain most of the variation in their early age studies, which is similar to the results of this study.
This research was conducted to compare different models based on predictions of genetic parameters and to show whether maternal effects could better explain the genetic effects on BW and WW traits. Overestimation of phenotypic variation, in other words, low total heritability, may be due to the low litter sizes, inconsistency in records, and poor maintenance, feeding, and herd management in the farms. It is thought that by improving these factors, the reliability in estimations would be increased.
Consequently, the maternal effect should be taken into consideration for the purpose of improving accuracy in parameter estimations and therefore increasing the success of breeding programs.
SPSS software (version 20.0) is available at: https://www.ibm.com/analytics/spss-statistics-software, last access: 14 March 2021 (IBM, 2020) and WOMBAT at: http://didgeridoo.une.edu.au/km/wombat.php, last access: 14 March 2021.
The data can be requested with written permission from the Republic of Turkey, General Directorate of Agricultural Research and Policies of the Ministry of Food, Agriculture and Livestock (https://www.tarimorman.gov.tr/TAGEM , last access: 14 March 2021).
The supplement related to this article is available online at: https://doi.org/10.5194/aab-65-121-2022-supplement.
All authors made substantial contributions to the study. HH performed project administration, supervision, conceptualization, data curation, and writing of the original draft. ÇT performed the formal analysis, methodology, and writing review and editing. EY collected the data. All authors read and approved the final article.
The contact author has declared that neither they nor their co-authors have any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This study was carried out by using the data obtained from the Awassi Small Ruminant Sub-Project (80İVE2012-01) of Public Animal Breeding National Project under the coordination of the Republic of Turkey, General Directorate of Agricultural Research and Policies of the Ministry of Food, Agriculture and Livestock.
This paper was edited by Antke-Elsabe Freifrau von Tiele-Winckler and reviewed by Zafer ULUTAŞ, Xingbo Zhao, and one anonymous referee.
Akaike, H.: Information theory as an extension of the maximum likelihood principle, 2nd Int. Symp. on Information Theory, Akademiai Kiado, Budapest, Hungary, 1973.
Aktaş, A. H. and Doğan, Ş.: Effect of live weight and age of Akkaraman ewes at mating on multiple birth rate, growth traits, and survival rate of lambs, Turk. J. Vet. Anim. Sci., 38, 176–182, https://doi.org/10.3906/vet-1301-10, 2014.
Çelikeloğlu, K. and Tekerli, M.: The Importance of Maternal Effect in Sheep Breeding, J. Bahri Dagdas Animal Research, 1–2, 29–32, ISSN 2148-3213, 2014.
Falconer, D. S. and Mackay, T. F. C.: Introduction to Quantitative Genetics, Longmans Green, Harlow, Essex, United Kingdom, ISBN 10 058224302, ISBN 13 9780582243026, 1996.
Felsenstein, J.: Evolutionary trees from DNA sequences: A maximum likelihood approach, J. Mol. Evol., 17, 368–376, 1981.
Galal, S., Gürsoy, O., and Shaat, I.: Awassi sheep as a genetic resource and efforts for their genetic improvement, Small Ruminant Res., 79, 99–108, 2008.
Gholizadeh, M., Rahimi Mianji, G., Hashemi, M., and Hafezian, H.: Genetic parameter estimates for birth and weaning weights in Raeini goats, Czech J. Anim. Sci., 55, 30–36, 2010.
Gowane, G. R., Chopra, A., Prakash, V., and Arora, A. L.: Estimates of (co)variance components and genetic parameters for body weights and first greasy fleece weight in Malpura sheep, Livest. Sci., 131, 94–101, 2010.
Gutierrez, J. P., Canon, J., and Goyache, F.: Estimation of direct and maternal genetic parameters for pre-weaning traits in the Asturiana de los Valles beef cattle breed through animal and sire models, J. Anim. Breed. Genet., 114, 261–266, https://doi.org/10.1111/j.1439-0388.1997.tb00511.x, 1997.
Hanrahan, J. P.: Maternal effects and selection response with an application to sheep data, Anim. Prod., 22, 359–369, 1976.
Heydarpour, M., Schaeffer, L. R., and Yazdi, M. H.: Influence of population structure on estimates of direct and maternal parameters, J. Anim. Breed. Genet., 125, 89–99, 2008.
Huelsenbeck, J. P. and Crandall, K. A.: Phylogeny estimation and hypothesis testing using maximum likelihood, Annu. Rev. Ecol. Syst., 28, 437–466, 1997.
IBM: SPSS software (version 20.0), https://www.ibm.com/analitics/spss-statistic-software, 2020.
Jafaroghli, M., Rashidi, A., Mokhtari, M., and Shadparvar, A.: (Co)Variance components and genetic parameter estimates for growth traits in Moghani sheep, Small Ruminant Res., 91, 170–177, 2010.
Jalil-Sarghale, A., Kholghi, M., Shahrebabak, M. M., Shahrebabak, H. M., Mohammadi, H., and Abdollahi-Arpanahi, R.: Model Comparisons and Genetic Parameter Estimates of Growth Traits in Baluchi Sheep, Slovak J. Anim. Sci., 47, 12–18, 2014.
Jawasreh, K., Ismail, Z. B., Iya, F., Castañeda-Bustos, V. J., and Valencia-Posadas, M.: Genetic parameter estimation for pre-weaning growth traits in Jordan Awassi sheep, Veterinary World, 11, 254–258, 2018.
Jawasreh, K. I. Z. and Khasawneh, A. Z.: Studies of some economic characteristic on Awassi lambs in Jordan, Egyptian J. Sheep Goat Sci., 2, 101–110, 2007.
Jawasreh, K. I. Z., Awawdeh, F. T., Al-Khasawneh, A. Z., Shdaifat, B., Al-Shboul, H., and Al-Hamed, B.: The effect of some placental factors in birth weight of Awassi lambs, Res. J. Anim. Vet. Sci., 4, 5–8, 2009.
Ligda, C., Gabriilidis, G., Papodopoulos, T., and Georgoudis, A.: Investigation of direct and maternal genetic effects on birth and weaning weight of Chios lambs, Livest. Prod. Sci., 67, 75–80, 2000.
López-Romero, P. and Carabano, M. J.: Comparing alternative random regression models to analyse first-lactation daily milk yield data in Holstein-Friesian cattle, Livest. Prod. Sci., 82, 81–96, 2003.
Maniatis, G., Demiris, N., Kranis, A., Banos, G., and Kominakis, A.: Model comparison and estimation of genetic parameters for body weight in commercial broilers, Can. J. Anim. Sci., 93, 67–77, 2013.
Meyer, K.: Estimating variances and covariances for multivariate animal models by Restricted Maximum Likelihood, Genet. Sel. Evol., 23, 67–78, 1991.
Meyer, K.: Bias and sampling covariances of estimates of variance components due to maternal effects, Genet. Sel. Evol., 24, 487–509, 1992.
Meyer, K.: WOMBAT, A tool for mixed model analyses in quantitative genetics by restricted maximum likelihood (REML), J. Zhejiang Univ. Sci. B., ISSN 1673-1581, ISSN 1862-1783, 8, 815–821, 2007.
Meyer, K. and Kirkpatrick, M.: Restricted maximum likelihood estimation of genetic principal components and smoothed covariance matrices, Genet. Sel. Evol., 37, 1–30, 2005.
Miraei-Ashtiani, Sr., Seyedalian, S., and Shahrbabak, M. M.: Variance components and heritabilities for body weight traits in Sangsari sheep, using univariate and multivariate animal models, Small Ruminant Res., 73, 109–114, 2007.
Mohammadabadi, M. R. and Sattayimokhtari, R.: Estimation of (co)variance components of ewe productivity traits in Kermani sheep, Slovak J. Anim. Sci., 46, 45–51, 2013.
Mohammadi, H., Shahrebabak, M. M., Vatankhah, M., and Shahrebabak, H. M.: Direct and maternal (co)variance components, genetic parameters, and annual trends for growth traits of Makooei sheep in Iran, Trop. Anim. Health Pro., 45, 185–191, 2013a.
Mohammadi, H., Shahrebabak, M. M., Shahrebabak, H. M., Bahrami, A., and Dorostkar, M.: Model comparisons and genetic parameter estimates of growth and the Kleiber ratio in Shal sheep, Arch. Anim. Breed., 56, 264–275, https://doi.org/10.7482/0003-9438-56-026, 2013b.
Mohammadi, H., Shahrebabak, M. M., Vatankhah, M., and Shahrebabak, H. M.: Direct and maternal (co)variance components, genetic parameters, and annual trends for growth traits of Makooei sheep in Iran, Trop. Anim. Health Pro., 45, 185–191, 2013c.
Prakash, V., Gowane, G. R., and Misra, S. S.: Application of WOMBAT for Analysis of maternal effects and SNP effect data analysis, 2016.
Pham, H.: A New Criterion for Model Selection, Mathematics, 7, 1215, 2019.
Rashidi, A., Mokhtari, M. S., Esmailizadeh, A. K., and Asadi Fozi, M.: Genetic analysis of ewe productivity traits in Moghani sheep, Small Ruminant Res., 96, 11–15, 2011.
Saatçi, M., Dewi, I. A., and Ulutas, Z.: Variance components due to direct and maternal effects and estimation of breeding values for 12 week weight of Welsh Mountain lambs, J. Anim. Sci., 69, 345–352, 1999.
Searle, S. R., Casella, G., and McCulloch, C. E.: Variance Components, John Wiley and Sons, New York, ISBN-I 3 978-0-470-00959-8, 2006.
Schwarz, G.: Estimating the dimension of a model, Ann. Stat., 6, 461–464, https://doi.org/10.1214/aos/1176344136, 1978.
Şireli, H. D., Vural, M. E., Karatas, A., Akca, N., Koncagül, S., and Tekel, N.: Birth and weaning weights of Awassi lambs raised in the GAP International Agricultural Research and Training Center, Ankara Univ. Vet. Fak., 62, 139–145, 2015.
Supakorn, C., Pralomkarn, W., and Anothaisinthawee, S.: Estimation of genetic parameters and genetic trends for weight and body measurements at birth in sheep populations in Thailand, Songklanakarin J. Sci. Technol., 35, 1–10, 2013.
Szwaczkowski, T., Wojtowski, J., Stanislawska, E., and Gut, A.: Estimates of maternal genetic and permanent environmental effects in sheep, Arch. Tierzucht, 49, 186–192, 2006.
Tesema, Z., Alemayehu, K., Getachew, T., Kebede, D., Deribe, B., Taye, M., Tilahun, M., Lakew, M., Kefale, A., Belayneh, N., Zegeye, A., and Yizengaw, L.: Estimation of genetic parameters for growth traits and Kleiber ratios in Boer x Central Highland goat, Trop. Anim. Health Pro., 52, 3195–3205, 2020.
TUIK: https://data.tuik.gov.tr/Kategori/GetKategori?p=Tarim-111, 2021.
Vatankhah, M. and Talebi, M. A.: Heritability estimates and correlations between production and reproductive traits in Lori-Bakhtiari sheep in Iran, S. Afr. J. Anim. Sci., 38, 110–118, 2008a.
Vatankhah, M., Talebi, M. A., and Edriss, M. A.: Estimation of genetic parameters for reproductive traits in LoriBakhtiari sheep, Small Ruminant Res., 74, 216–220, 2008b.
Vatankhah, M., Moradi, M., Nejati-Javaremi, A., MireaeiAshtiani, S. R., and Vaez-Torshizi, R.: A review of sheep breeding in Iran, in: Proc. 1st Congress on Animal and Aquatic Sciences, Tehran, Iran, 591–597, 2004.
Willham, R. L.: The role of maternal effects in animal breeding. III. Biometrical aspects of maternal effects in animals, J. Anim. Sci., 35, 1288–1293, 1972.
Yavarifard, R., Ghavi, H. Z., and Shadparvar, A. A.: Estimation of genetic parameters for reproductive traits in Mehraban sheep, Czech J. Anim. Sci., 60, 281–288, https://doi.org/10.17221/8242-CJAS, 2015.
Zhang, C. Y., Zhang, Y., Xu, D. Q., Li, X., Su, J., and Yang, L. G.: Genetic and phenotypic parameter estimates for growth traits in Boer goat, Livest. Sci., 124, 66–71, 2009.